Różnice hemodynamiczne między prawą a lewą komorą | Revista Española de Cardiología
Lewa komora jest pompą ciśnieniową, która wysyła bolus wyrzutowy przy dużych oporach obwodowych, podczas gdy prawa komora jest pompą objętościową, która wysyła ten sam bolus wyrzutowy przy małych oporach obwodowych.
Tak jak lewa komora była szeroko badana, tak w przypadku prawej komory przyjęto, że z wyjątkiem wyraźnie niższego ciśnienia skurczowego i nieco niższej frakcji wyrzutowej (55%), jej funkcja jest identyczna z zachowaniem lewej komory.
Artykuł opublikowany w tym numerze Revista Española de Cardiología przez Juan C. Grignola i wsp. (str. 37-42)1 , badający ciśnienia w prawej komorze, tętnicy płucnej, lewej komorze, aorcie, przepływ płucny i średnice komór za pomocą sonomrometrii w sercach owiec, pokazuje, jak niedokładne jest to twierdzenie. Według tych autorów prawa komora ma wydłużoną fazę wyrzutu i brak w niej izowolumetrycznej fazy relaksacji.
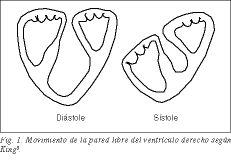
Prawa komora, utworzona przez pętlę podstawną z tętnicy płucnej, zgodnie z koncepcją Torrenta 1 , obejmująca pętlę koniuszkową lewej komory, jest rozszerzeniem lewej komory, sprzężonym z normalnym funkcjonowaniem lewej komory i przez nie uwarunkowanym. Jego duża powierzchnia w kształcie półksiężyca pozwala na dostarczanie dużych ilości krwi przy niskich oporach płucnych i przy niskich kosztach. Z licznych starych doświadczeń wynika, że po usunięciu wolnej ściany prawej komory u psów nie pojawiają się objawy niewydolności prawego serca, ponieważ wypukła przegroda lewej komory aktywnie współpracuje przy wyrzucie z prawej komory 2-5 . Skrócenie podłużne pasma tworzącego wolną ścianę prawej komory powoduje skurczowe obniżenie pierścienia trójdzielnego i zgniatanie wolnej ściany nad przegrodą jak miech. Już King, lekarz z Guy’s Hospital w Londynie w 1837 roku, uważał, że przegroda jest ścianą litą, podczas gdy wolna ściana byłaby ścianą ruchomą lub plastyczną (ryc. 1) 6 . Przegroda, wybrzuszając się w kierunku prawej jamy, współpracuje z wolną ścianą prawej komory w efekcie działania miecha.
Ale prawa komora składa się z dwóch części: części napływowej lub zatoki, która zaczyna się na zastawce trójdzielnej i części wypływowej lub infundibulum, która kończy się na zastawce płucnej. W badaniach Grignoli i wsp. zaobserwowano, że wyrzut z komory składa się z dwóch etapów, wczesnego i późnego. Jest to w pełni zgodne z obserwacjami anatomicznymi, embriologicznymi, anatomii porównawczej i elektrofizjologicznymi, które sugerują, że część napływowa i odpływowa prawej komory są różnymi komorami, które ponadto pojawiły się w różnych okresach ewolucji.
Infundibulum jest umięśnioną częścią drogi odpływu serca, która pojawia się filogenetycznie już u najprymitywniejszych chordates, takich jak bezszczękowe cyklostomy (minóg) w okresie syluru ery paleozoicznej, 435 milionów lat temu 7 . W dewonie, 400 milionów lat temu, rekiny z podklasy elasmobranchs mają w pełni umięśnione infundibulum (bulbus cordis) z okrężnymi włóknami 8 . U salamandry (urodelo), współczesnego płaza z epoki Mississippian okresu karbońskiego sprzed 345 milionów lat, infundibulum jest tak rozwinięte, że przypomina niektóre przypadki stenozy płucnej.
Napływowa część prawej komory, z drugiej strony, jest znacznie nowsza, zaczyna pojawiać się u gadów z epoki Pennsylvanian okresu karbońskiego, również ery paleozoicznej, około 275 milionów lat temu, kiedy to pojawia się początkowa koniuszkowa przegroda międzykomorowa, prawdopodobnie jako przystosowanie do rozdzielenia krążenia systemowego i płucnego podczas oddychania powietrzem 9 . U krokodyla, po raz pierwszy w historii ewolucji, oba krążenia żylne i tętnicze są obecnie całkowicie oddzielone, z nienaruszoną przegrodą międzykomorową, a infundibulum jest włączone do prawej komory, chociaż pełne włączenie występuje u ptaków i ssaków 10 . Rozwój prawej komory może być uważany za dehiscencję lewej komory, według Torrenta Guaspa (komunikacja osobista) (ryc. 2).

Ontogenetycznie infundibulum ma również bardzo wczesny rozwój embriologiczny, pojawiając się w stadium prostej rurki sercowej w 23-24 dniu rozwoju. Kiedy przewód jest zakrzywiony i tworzy się prawa pętla wypukła, zatoka prawej komory powstaje przez rozszerzenie się części ogonowej bulbus cordis, która tworzy infundibulum, podczas gdy przegroda międzykomorowa „rośnie” biernie przez rozszerzenie się dwóch jam komorowych po każdej stronie (dzień 27) 11 .
Kilka obserwacji anatomicznych wskazuje również, że zatoka i infundibulum prawej komory stanowią odrębne elementy. Dwukomorowa prawa komora jest zmianą charakteryzującą się postępującym zwężeniem napływu do pęczka Hisa, prowadzącym do niedrożności pomiędzy zatoką a pęczkiem Hisa. W dwuwlotowej lewej komorze, najczęstszej formie pojedynczej komory, zatoka prawej komory jest nieobecna, a infundibulum jest zawsze obecne.
Badania elektrofizjologiczne wskazują również, że aktywacja infundibulum następuje stosunkowo późno w skurczu i ta część serca aktywuje się jako ostatnia, co może skutkować asynchronicznym skurczem i rozkurczem części napływowej i odpływowej prawej komory 12 .
Ginés i wsp. wykazują u owiec, że w skurczu, w prawej komorze maksymalny wyrzut jest osiągany wcześnie, a koniec wyrzutu jest znacznie większy niż w lewej komorze. Innymi słowy, skurcz prawej komory jest skurczem perystaltycznym, który przebiega od części napływowej do odpływowej (infundibulum) prawej komory. Ten fakt, który został zasugerowany u psa i wykazany w tej pracy u owiec, został teraz udowodniony również u człowieka. Za pomocą jednoczesnej akustycznej echokardiografii kwantylowej, fonokardiografii i rezonansu magnetycznego potwierdzono istnienie regionalnych różnic pomiędzy częścią napływową i napływową prawej komory, które polegają na tym, że część napływowa rozszerza się podczas skurczu części napływowej, a kurczy się po komponencie aortalnej drugiego tonu 13 .
Drugim ważnym faktem tej pracy jest wykazanie nieistnienia izowolumetrycznej fazy relaksacji w prawej komorze, a naszym zdaniem również izowolumetrycznej fazy skurczu.
Mechanizm, dzięki któremu komory serca wypełniają się w rozkurczu, był szeroko dyskutowany w całej historii medycyny. Erasistratus w IV wieku p.n.e. uważał, że komora serca działa jak pompa ssąca podczas rozkurczu. Jednak Harvey, który zmienił obowiązujące od czasów Galena pojęcia skurczu i rozkurczu, w swoim „Exercitatio anatomica de motu cordis et sanguinis in animalibus” stwierdził: „…sposób, w jaki krew dostaje się do komór, jest oczywisty: nie dlatego, że serce (komora) wciąga ją, ponieważ się rozszerza, lecz dlatego, że jest do niej wrzucana przez puls (skurcz) małżowin usznych” 14 . Później Wiggers, biorąc pod uwagę, że napełnianie następuje głównie na początku rozkurczu, doszedł do wniosku, że napełnianie zależy od różnicy ciśnień między przedsionkiem a komorą pod koniec izometrycznej fazy relaksacji 15 . Po latach jednak był w stanie stwierdzić, że „komory mają elastyczny odrzut, który pozwala im wypełnić się prawie całkowicie przed skurczem przedsionków” 16 .
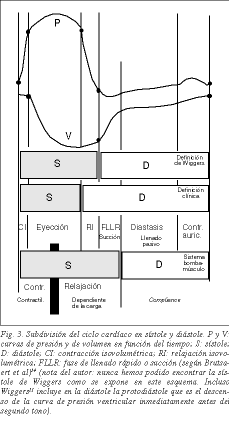
Współczesna koncepcja aktywnego rozkurczu ma swój początek u Brechera, który na sugestię Torrenta Guaspa zdołał wykazać istnienie ssania komorowego17 . Kilka lat temu Dirk Brutsaert i wsp.18 zaproponowali, aby skurcz rozumieć raczej z konceptualnego niż fenomenologicznego punktu widzenia i aby obejmował on relaksację serca z izowolumetrycznymi fazami relaksacji oraz fazę szybkiego napełniania komór (ryc. 3). Rzeczywiście, relaksacja, definiowana jako proces, w którym serce powraca do konfiguracji sprzed skurczu, jest procesem aktywnym. Zużycie ATP jest głównie wykorzystywane w procesie relaksacji w podwójnej ścieżce: zużycie ATP przez ATPazę z głów miozyny w celu odłączenia miozyny od aktyny oraz przez SERCAs w celu katapultowania Ca+ z troponiny C do siateczki sarkoplazmatycznej, oba procesy sygnalizują początek relaksacji. Tak więc jednym z kamieni milowych ostatnich lat w mechanizmie skurczu mięśni jest wiedza, że aktywnym procesem maksymalnego zużycia energii jest relaksacja 19,20 . Dlatego cykl serca można nadal dzielić na skurcz i rozkurcz, jak to tradycyjnie czyniono od czasów Harveya, ale ze świadomością, że pierwsza część rozkurczu jest aktywna, tzn. jest to izowolumetryczna faza relaksacji i faza szybkiego napełniania lub zasysania komór.
W mechanizm zasysania zaangażowanych jest wiele sił, z których niektóre muszą być bardzo silne, aby wyjaśnić przypadek Robertsa i wsp.21 , w którym rana postrzałowa prawej komory spowodowała tak gwałtowne zasysanie, że doszło do inwazji lewego przedsionka do lewej komory. Bierne sprężyste zwijanie się spirali komory podczas jej rozkurczu może mieć zasadnicze znaczenie, choć efekt „rękawa doprowadzającego krew” w tętnicach wieńcowych podczas ich napełniania w rozkurczu oraz prawo Laplace’a, gdy ściana komory rozrzedza się w rozkurczu i zwiększa się jej promień, również mogą odgrywać pewną rolę. Ostatnio Torrent Guasp zasugerował, że serce funkcjonuje jak tłok, w którym wyrzut odbywa się przez teleskopowe wydłużenie pętli podstawnej nad pętlą apexiańską (opadanie tłoka w dół cylindra), a ssanie byłoby wydłużeniem-poszerzeniem komór przez wyprostowanie przegrody ciągnącej pętlę podstawną (wznoszenie się tłoka) 2 .
Ale to się dzieje w lewej komorze. Dlaczego nie ma relaksacji izowolumetrycznej w prawej komorze i dlatego nie ma ssania? Oczywiste jest, że jeśli w prawej komorze występuje skurcz, to musi nastąpić relaksacja, a jeśli występuje wyrzut, to musi nastąpić napełnianie. Jedną z możliwości jest to, że krzywe objętościowo-ciśnieniowe prawej komory w kształcie trójkąta rejestrują dwa odrębne i jednoczesne zdarzenia: skurcz części napływowej i napełnianie znacznie większej części napływowej. Istnieją co najmniej dwie prace, które potwierdzają ten pogląd. March i wsp.21 zasugerowali, że prawa komora składa się z dwóch niezależnych komór, a Pouleur i wsp.22 potwierdzili, że wyrzut z prawej komory jest kontynuowany podczas relaksacji wolnej ściany, co Shaver i wsp. postulowali już wcześniej z osłuchowego punktu widzenia i określili jako „uduszenie”, aby wyjaśnić opóźnienie komponentu płucnego drugiego tonu.
Inna możliwość byłaby taka, że niska impedancja drzewa płucnego nie wymaga fazy ssania, a prawe serce wypełnia się przez prosty spadek ciśnienia przedsionkowo-komorowego. Za tym prostszym poglądem przemawiałoby to, że jeśli nie ma izowolumetrycznej fazy relaksacji, nie ma też potrzeby istnienia izowolumetrycznej fazy skurczu, również nieobecnej w krzywych objętość-ciśnienie Grignoli i wsp. i pojawiającej się za to w nadciśnieniu płucnym, gdzie krzywe objętość-ciśnienie stają się podobne do tych po lewej stronie.
Czy to wysokie obciążenie następcze wymaga fazy izowolumetrycznego skurczu i izowolumetrycznego rozkurczu, aby wywołać ssanie?
W każdym razie różnice między trójkątnym kształtem krzywych objętość-ciśnienie w prawej komorze a kwadratowymi krzywymi objętość-ciśnienie w lewej komorze są fascynującym i obiektywnym faktem, którego hemodynamiczna interpretacja wymaga dalszych rozważań i badań.