Hemodynamiska skillnader mellan höger och vänster kammare | Revista Española de Cardiología
Vänster kammare är en tryckpump som skickar en utstötningsbolus mot höga perifera motstånd, medan höger kammare är en volympump som skickar samma utstötningsbolus mot låga perifera motstånd.
På samma sätt som den vänstra kammaren har studerats ingående har man antagit att den högra kammaren, med undantag för det markant lägre systoliska trycket och den något lägre ejektionsfraktionen (55 %), har samma funktion som den vänstra kammaren.
Den artikel som publiceras i detta nummer av Revista Española de Cardiología av Juan C. Grignola et al (s. 37-42)1 , där trycket i höger kammare, lungartären, vänster kammare, aorta, lungflödet och kammardiametern undersöks med hjälp av sonomikrometri i fårhjärtan, visar hur felaktigt detta påstående är. Enligt dessa författare har höger kammare en förlängd ejektionsfas och saknar isovolumisk relaxationsfas.
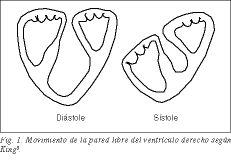
Den högra kammaren, som bildas av den basala slingan från lungartären, enligt Torrents koncept 1 , och som omfattar den apikala slingan från den vänstra kammaren, är en utvidgning av den vänstra kammaren, kopplad till och betingad av den vänstra kammarens normala funktion. Dess stora halvmåneformade yta gör att den kan leverera stora mängder blod mot låga lungmotstånd och till låg kostnad. Det finns många gamla erfarenheter som visar att när man tar bort den fria väggen i den högra kammaren hos hundar uppträder inga tecken på höger hjärtsvikt, eftersom den vänstra kammarens konvexa septum aktivt medverkar till utstötningen av den högra kammaren 2-5 . Den longitudinella förkortningen av det band som bildar högerkammarens fria vägg resulterar i systolisk sänkning av trikuspidalringen och krossning av den fria väggen över septum som en bälg. Redan King, en läkare vid Guy’s Hospital i London 1837, hade betraktat septum som en fast vägg, medan den fria väggen skulle vara en rörlig eller duktil vägg (fig. 1) 6 . Genom att septum böljar ut mot den högra kaviteten samarbetar det med den fria väggen i den högra ventrikeln i bälgeffekten.
Men den högra kammaren består av två delar: en inflödesdel eller sinus som börjar vid trikuspidalklaffen och en utflödesdel eller infundibulum som slutar vid lungklaffen. I Grignola et al:s resultat observeras att den ventrikulära utstötningen består av två stadier, tidigt och sent. Detta stämmer helt överens med anatomiska, embryologiska, komparativa anatomiska och elektrofysiologiska observationer, som tyder på att inflödes- och utflödesdelarna av höger kammare är olika kamrar som dessutom uppstod under olika utvecklingsperioder.
Infundibulum är en muskulös del av hjärtats utflödeskanal som förekommer fylogenetiskt så tidigt som de mest primitiva chordaterna, t.ex. de käklösa cyklostomerna (nejonögon) under silurperioden i paleozoikum, för 435 miljoner år sedan 7 . I Devon, för 400 miljoner år sedan, hade hajar i underklassen elasmobrancher ett fullt muskulärt infundibulum (bulbus cordis) med cirkulära fibrer 8 . Hos salamandern (urodelo), en modern amfibie från den Mississippiska epoken i Karbon för 345 miljoner år sedan, är infundibulum så utvecklat att det påminner om vissa fall av lungstenos.
Den inkommande delen av den högra kammaren är å andra sidan mycket nyare och börjar uppträda hos reptiler från den pennsylvanska epoken i karbonperioden, även den från paleozoikum, för cirka 275 miljoner år sedan, när den begynnande apikala interventrikulära septumet uppstår, möjligen som en anpassning till separationen av den systemiska och lungcirkulationen när man andas luft 9 . Hos krokodilen är de två venösa och arteriella cirkulationerna nu för första gången i evolutionens historia helt separerade med en intakt interventrikulär septum, och infundibulum är inkorporerat i den högra ventrikeln, även om fullständig inkorporering förekommer hos fåglar och däggdjur 10 . Utvecklingen av den högra ventrikeln kan betraktas som en dehiscens av den vänstra ventrikeln, enligt Torrent Guasp (personlig kommunikation) (fig. 2).

Ontogenetiskt sett har infundibulum också en mycket tidig embryologisk utveckling och uppträder i det raka hjärtrörsstadiet vid 23-24 utvecklingsdagar. När röret är böjt och den högra konvexa slingan bildas, bildas den högra ventrikelsinus genom expansion av den kaudala delen av bulbus cordis, som bildar infundibulum, medan den interventrikulära septum ”växer” passivt genom expansion av de två ventrikulära kaviteterna på vardera sidan (dag 27) 11 .
Flera anatomiska observationer visar också att den högra ventrikelns sinus och infundibulum är skilda komponenter. Dubbelkammarhögerkammaren är en skada som kännetecknas av progressiv förträngning av infundibulums inflöde, vilket leder till obstruktion mellan sinus och infundibulum. I vänster ventrikel med dubbla inlopp, den vanligaste formen av enkel ventrikel, saknas den högra ventrikelns sinus och infundibulum är alltid närvarande i stället.
Elektrofysiologiska studier visar också att aktiveringen av infundibulum sker relativt sent i systolen och att denna del av hjärtat är den sista som aktiveras, vilket kan resultera i asynkron kontraktion och relaxation av inflödes- och utflödesdelarna av höger kammare 12 .
Ginés et al. visar på får att i systole, i den högra kammaren, uppnås maximal ejektion tidigt och att slutet av ejektionen är betydligt större än i den vänstra kammaren. Med andra ord är sammandragningen av höger kammare en peristaltisk sammandragning som går från inflödet till utflödesdelen (infundibulum) av höger kammare. Detta faktum, som föreslagits hos hundar och som i detta arbete demonstreras hos får, har nu också bevisats hos människor. Genom samtidig akustisk kvantifierad ekokardiografi, fonokardiografi och magnetisk resonanstomografi har det bekräftats att det finns regionala skillnader mellan högerkammarens inflödesdel och infundibulum, så att infundibulum expanderar när inflödesdelen drar ihop sig och drar ihop sig efter aortakomponenten i den andra tonen 13 .
Det andra viktiga resultatet av detta arbete är beviset för att det inte finns någon isovolumetrisk relaxationsfas i höger kammare, och enligt vår åsikt inte heller någon isovolumetrisk kontraktionsfas.
Mekanismen genom vilken ventriklarna fylls i diastole har debatterats flitigt under medicinens historia. Erasistratus på 400-talet f.Kr. trodde att ventrikeln fungerade som en sugpump under diastolen. Harvey, som ändrade begreppen systole och diastole sedan Galen, sade i sin ”Exercitatio anatomica de motu cordis et sanguinis in animalibus”: ”… det är uppenbart hur blodet kommer in i ventriklarna: inte för att hjärtat (ventrikeln) drar in det genom att det utvidgas, utan för att det kastas in i det genom pulsen (sammandragningen) av auriklarna” 14 . Senare kom Wiggers, med hänsyn till att fyllningen huvudsakligen sker i början av diastolen, fram till slutsatsen att fyllningen beror på tryckskillnaderna mellan förmak och kammare i slutet av den isometriska relaxationsfasen 15 . Flera år senare kunde han dock konstatera att ”ventriklarna har en elastisk rekyl som gör att de kan fyllas nästan helt innan förmaksinkontraktionen” 16 .
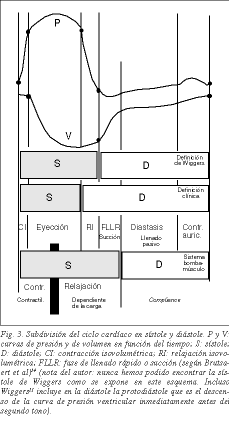
Det moderna begreppet aktiv diastole börjar med Brecher som, på förslag av Torrent Guasp, kunde påvisa förekomsten av ventrikelsug17 . För flera år sedan föreslog Dirk Brutsaert et al18 att systole bör förstås ur en konceptuell snarare än fenomenologisk synvinkel och att den bör inkludera kardiell relaxation med isovolumetriska relaxationsfaser och den snabba kammarfyllnadsfasen (fig. 3). Relaxation, som definieras som den process där hjärtat återgår till sin konfiguration före sammandragning, är faktiskt en aktiv process. ATP-förbrukningen används främst i relaxationsprocessen i en dubbel väg: ATP-förbrukning av ATPas från myosinhuvuden för att lossa myosin från aktin och av SERCAs för att katapultera Ca+ från troponin C till det sarkoplasmatiska retikulumet, båda processerna signalerar att relaxation inleds. En av de senaste årens milstolpar inom mekanismen för muskelkontraktion är därför kunskapen om att den aktiva processen för maximal energiförbrukning är avslappning 19,20 . Hjärtcykeln kan därför fortfarande delas in i systole och diastole som traditionellt gjorts sedan Harvey, men med vetskapen att den första delen av diastolen är aktiv, dvs. den isovolumetriska relaxationsfasen och den snabba kammarfyllnads- eller sugfasen.
Många krafter är inblandade i sugmekanismen, och några av dem måste vara mycket starka för att förklara fallet med Roberts et al 21 , där ett skottskada i den högra kammaren orsakade ett så våldsamt sug att den vänstra förmaksbilagan invaginerades in i den vänstra kammaren. Den passiva elastiska rekylen hos den ventrikulära spiralen när den slappnar av kan vara grundläggande, även om effekten av kranskärlens ”blodförsörjningsmanschett” när de fylls i diastole och Laplaces lag när den ventrikulära väggen tunnas ut i diastole och radien ökar också kan spela en roll. Torrent Guasp har nyligen föreslagit att hjärtat fungerar som en kolv där utstötning sker genom att den basala slingan teleskoperas över den apexianska slingan (kolven sjunker ner i cylindern) och sugning skulle vara en förlängning och breddning av ventriklarna genom att septumet rätas ut och drar den basala slingan (kolvens uppstigning) 2 .
Men detta sker i vänster kammare. Varför sker ingen isovolumetrisk relaxation i höger kammare och därmed ingen sugning? Det är uppenbart att om det sker en sammandragning i höger kammare måste det ske en avslappning, och om det sker en utstötning måste det ske en fyllning. En möjlighet är att de triangelformade högerkammarvolym-tryckkurvorna registrerar två distinkta och samtidiga händelser: kontraktion av infundibulum och fyllning av den mycket större inflödesdelen. Det finns minst två artiklar som stöder denna åsikt. March et al 21 föreslog att högerkammaren består av två oberoende kamrar och Pouleur et al 22 bekräftade att högerkammarens utstötning fortsätter när den fria väggen slappnar av, vilket Shaver et al redan hade förespråkat ur auskultationssynpunkt och betecknat som ”strangulering” för att förklara fördröjningen av den pulmonella komponenten i den andra tonen.
Den andra möjligheten är att den låga impedansen hos lungträdet inte kräver någon sugfas och att högerhjärtat fylls genom enkel atrioventrikulär tryckminskning. För detta enklare synsätt talar att om det inte finns någon isovolumetrisk relaxationsfas finns det inte heller något behov av en isovolumetrisk kontraktionsfas, som också saknas i Grignola et al:s volymtryckskurvor och som i stället uppträder vid pulmonell hypertoni där volymtryckskurvorna liknar dem till vänster.
Är det den höga efterbelastningen som kräver en fas av isovolumetrisk kontraktion och isovolumetrisk relaxation för att orsaka sugning?
I vilket fall som helst är skillnaderna mellan den triangulära formen på höger kammarvolym-tryckkurvorna och de kvadratiska vänsterkammarvolym-tryckkurvorna ett fascinerande och objektivt faktum, vars hemodynamiska tolkning kräver ytterligare eftertanke och studier.