Diferencias hemodinámicas entre el ventrículo derecho e izquierdo | Revista Española de Cardiología
El ventrículo izquierdo es una bomba de presión que envía un bolo de eyección contra altas resistencias periféricas, mientras que el ventrículo derecho es una bomba de volumen que manda el mismo bolo de eyección contra bajas resistencias periféricas.
Así como se ha estudiado extensamente el ventrículo izquierdo, en el ventrículo derecho se ha asumido que, excepto por la presión sistólica que es notablemente inferior y por la fracción de eyección que es ligeramente menor (55%), es idéntico en su función al comportamiento del ventrículo izquierdo.
El artículo publicado en este número de la Revista Española de Cardiología por Juan C. Grignola et al (pág. 37-42) 1 , estudiando las presiones del ventrículo derecho, arteria pulmonar, ventrículo izquierdo, aorta, flujo pulmonar y diámetros ventriculares por sonomicrometría, en corazones de oveja, demuestra cuán inexacto es este aserto. Según demuestran estos autores, el ventrículo derecho tiene una fase de eyección prolongada y carece de la fase de relajación isovolumé trica.
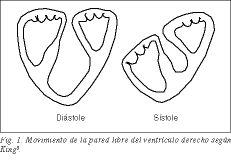
El ventrículo derecho formado por la lazada basal que parte de la arteria pulmonar, según el concepto de Torrent 1 , abrazando la lazada apical del ventrículo izquierdo, es una expansión del ventrículo izquierdo, acoplado y condicionado al normal funcionamiento de éste. Su gran superficie de forma semilunar le permite enviar grandes cantidades de sangre contra bajas resistencias pulmonares y a bajo coste. Hay numerosas experiencias antiguas que, eliminando la pared libre del ventrículo derecho en perros, no aparecen signos de insuficiencia cardíaca derecha, porque el septo convexo del ventrículo izquierdo colabora activamente en la eyección del ventrículo derecho 2-5 . El acortamiento longitudinal de la banda que forma la pared libre del ventrículo derecho da lugar al descenso sistólico del anillo tricúspide y el aplastamiento de la pared libre sobre el tabique como si fuera un fuelle. Ya King, un médico del Guy’s Hospital de Londres en 1837, había considerado el tabique como una pared sólida , mientras que la pared libre sería una pared móvil o dúctil (fig. 1) 6 . El septo, al abombarse hacia la cavidad derecha, colabora con la pared libre del ventrículo derecho en el efecto de fuelle.
Pero el ventrículo derecho consta de dos partes: una porción de entrada o seno que comienza en la válvula tricúspide y una porción de salida o infundíbulo que termina en la válvula pulmonar. En los hallazgos de Grignola et al se observa que la eyección del ventrículo consta de dos etapas, temprana y tardía. Este hecho está totalmente de acuerdo con observaciones anatómicas, embriológicas, de anatomía comparada y electrofisiológicas, que sugieren que la porción de entrada y la porción de salida del ventrículo derecho son cámaras diferentes que, además, aparecieron en períodos distintos de la evolución.
El infundíbulo es una porción muscularizada del tracto de salida del corazón que aperece filogenéticamente ya en los cordados más primitivos como los ciclóstomos sin mandíbula (la lamprea), en el período silúrico de la era paleozoica, hace 435 millones de años 7 . En el devónico, hace 400 millones de años, los tiburones de la subclase elasmobranquios tienen un infundíbulo (bulbus cordis) totalmente muscularizado con fibras circulares 8 . En la salamandra (urodelo), un anfibio moderno de la época misisipiana del período carbonífero hace 345 millones de años, el infundíbulo está tan desarrollado que recuerda algunos casos de estenosis pulmonar.
La porción de entrada del ventrículo derecho, en cambio, es mucho más reciente, comenzando a aparecer en los reptiles de la época pensilveniana del período carbonífero, también de la era paleozoica, hace unos 275 millones de años, cuando surge el incipiente septo interventricular apical, posiblemente como una adaptación a la separación de la circulación sistémica y pulmonar al respirar aire 9 . En el cocodrilo, por primera vez en la historia evolutiva, las dos circulaciones venosa y arterial están ya separadas por completo con un septo interventricular íntegro, y el infundíbulo está incorporado en el ventrículo derecho, aunque la incorporación total se hace en las aves y mamíferos 10 . El desarrollo del ventrículo derecho se puede considerar como una dehiscencia del ventrículo izquierdo, según Torrent Guasp (comunicación personal) (fig. 2).

Ontogénicamente el infundíbulo es también de desarrollo embriológico muy precoz, apareciendo ya en el estadio de tubo cardíaco recto a los 23-24 días de desarrollo. Cuando se incurva el tubo y se forma el asa de convexidad derecha se forma el seno del ventrículo derecho por expansión de la parte caudal del bulbus cordis, que forma el infundíbulo, al mismo tiempo que el septo interventricular «crece» pasivamente por expansión de las dos cavidades ventriculares a cada lado (día 27) 11 .
Varias observaciones anatómicas también demuestran que el seno y el infundíbulo del ventrículo derecho son componentes distintos. El ventrículo derecho de doble cámara es una lesión caracterizada por una estenosis progresiva de la entrada del infundíbulo que conduce a una obstrucción entre el seno y el infundíbulo. En el ventrículo izquierdo de doble entrada, la forma más común de ventrículo único, el seno del ventrículo derecho está ausente y en cambio existe siempre el infundíbulo.
Estudios electrofisiológicos indican también que la activación del infundíbulo ocurre relativamente tarde en la sístole y que esta parte del corazón es la última en activarse, lo que puede dar lugar a la contracción y relajación asincrónica de la porción de entrada y de salida del ventrículo derecho 12 .
Ginés et al demuestran en la oveja que en la sístole, en el ventrículo derecho, la eyección máxima se alcanza precozmente y que el fin de la eyección es significativamente mayor que en el ventrículo izquierdo. Es decir, que la contracción del ventrículo derecho es una contracción peristáltica que avanza de la porción de entrada a la porción de salida (infundíbulo) del ventrículo derecho. Este hecho, que se había sugerido en el perro y que se demuestra en este trabajo en ovejas, se acaba de comprobar también en el hombre. Por ecocardiografía cuantificada acústica, fonocardiografía y cine de resonancia magnética simultánea se ha confirmado que existen diferencias regionales entre la porción de entrada y el infundíbulo del ventrículo derecho, de modo que el infundíbulo se expande cuando se contrae la porción de entrada y se contrae después del componente aórtico del segundo tono 13 .
El segundo hecho importante de este trabajo es la demostración de la inexistencia de la fase de relajación isovolumétrica en el ventrículo derecho, y en nuestra opinión, también de la fase de contracción isovolumétrica.
El mecanismo por el que se llenan los ventrículos en la diástole se ha debatido ampliamente a lo largo de la historia de la medicina. Erasistrato en el siglo iv a.C. creyó que el ventrículo actuaba como una bomba de succión durante la diástole. Sin embargo, Harvey, que cambió los conceptos de sístole y diástole que se mantenían desde Galeno, decía en su «Exercitatio anatomica de motu cordis et sanguinis in animalibus»: «…resulta manifiesto la manera como la sangre entra a los ventrículos: no porque el corazón (ventrículo) la atraiga porque se dilate, sino porque es arrojada en su interior por el pulso (contraccion) de las aurículas» 14 . Posteriormente, Wiggers, teniendo en cuenta que el llenado se hace fundamentalmente al principio de la diástole, llegó a la conclusión de que el llenado dependía de las diferencias de presión entre aurícula y ventrículo al final de la fase de relajación isométrica 15 . Años después, sin embargo, pudo afirmar que «los ventrículos tienen un retroceso elástico que les permite llenarse prácticamente del todo antes de la contracción auricular» 16 .
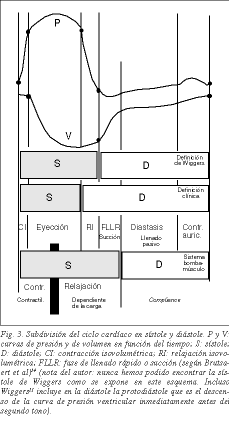
El concepto moderno de la diástole activa comienza con Brecher que, por sugerencia de Torrent Guasp, pudo demostrar la existencia de succión ventricular17 . Hace ya varios años, Dirk Brutsaert et al 18 habían propuesto que la sístole se debía entender desde el punto de vista más conceptual que fenomenológico y que debía incluir la relajación cardíaca con las fases de relajación isovolumétrica y la fase de llenado rápido ventricular (fig. 3). En efecto, la relajación, definida como el proceso en que el corazón retorna a su configuración precontráctil, es un proceso activo. El consumo de ATP se emplea fundamentalmente en el proceso de la relajación en una doble vía: el consumo de ATP por la ATPasa de las cabezas de miosina para despegar la miosina de la actina y por los SERCA para catapultar el Ca ++ de la troponina C al retículo sarcoplásmico, ambos procesos señalando el comienzo de la relajación. Así que uno de los hitos de los últimos años en el mecanismo de la contracción muscular es el conocimiento de que el proceso activo de máximo consumo energético es la relajación 19,20 . Por tanto, se puede seguir dividiendo el ciclo cardíaco en sístole y diástole como tradicionalmente se hacía desde Harvey, pero sabiendo que la primera parte de la diástole es activa, es decir, la fase de relajación isovolumétrica y la fase de llenado rápido ventricular o succión.
En el mecanismo de succión colaboran muchas fuerzas, alguna de las cuales tiene que ser muy potente para poder explicar el caso de Roberts et al 21 , en el que una herida de bala en el ventrículo derecho originó una succión tan violenta que invaginó la orejuela izquierda en el ventrículo izquierdo. El retroceso elástico pasivo del helicoide ventricular al relajarse puede ser fundamental, aunque también colaboran el efecto de «manga de riego» de las arterias coronarias al llenarse en diástole y la ley de Laplace al adelgazarse la pared ventricular en diástole y aumentar el radio. Recientemente, Torrent Guasp ha sugerido que el corazón funciona como un pistón en que la eyección se hace por el telescopaje de la lazada basal sobre la lazada apexiana (descenso del pistón por el cilindro) y la succión sería el alargamiento-ensanchamiento de los ventrículos por el enderezamiento del septo arrastrando la lazada basal (ascenso del pistón) 2 .
Pero esto ocurre en el ventrículo izquierdo. ¿Por qué no existe relajación isovolumétrica en el ventrículo derecho y, por lo tanto, succión? Evidentemente si en el ventrículo derecho hay contracción tiene que haber relajación y si hay eyección tiene que haber llenado. Una posibilidad podría ser que las curvas de volumen-presión del ventrículo derecho, de forma triangular, estuvieran registrando dos acontecimientos distintos y simultáneos: la contracción del infundíbulo y el llenado de la porción de entrada, que es mucho mayor. Hay, por lo menos, dos trabajos que apoyan este punto de vista. March et al 21 sugieren que el ventrículo derecho consta de dos cámaras independientes y Pouleur et al 22 confirmaron que la eyección del ventrículo derecho continúa cuando se está relajando la pared libre, lo que Shaver et al ya habían defendido desde el punto de vista de la auscultación y designaron como «estrangulación» para explicar el retraso del componente pulmonar del segundo tono.
La otra posibilidad sería que la baja impedancia del árbol pulmonar no necesitara fase de succión y el corazón derecho se llenara por simple desnivel de presión auriculoventricular. En favor de este punto de vista más sencillo estaría que si no hay fase de relajación isovolumétrica tampoco se necesita fase de contracción isovolumétrica, también inexistente en las curvas de volumen-presión de Grignola et al y en cambio aparece en la hipertensión pulmonar en que las curvas de volumen-presión se hacen semejantes a las izquierdas.
¿Es la elevada poscarga la que exige una fase de contracción isovolumétrica y de relajación isovolumétrica para originar la succión?
En cualquier caso, las diferencias entre la forma triangular de las curvas de volumen-presión del ventrículo derecho y las curvas cuadrangulares de volumen-presión del ventrículo izquierdo son un hecho fascinante y objetivo, cuya interpretación hemodinámica necesita posterior reflexión y estudio.