右心室と左心室の血行動態の違い|Revista Española de Cardiología
左心室は高い末梢抵抗に対して駆出ボーラスを送る圧力ポンプであり、右心室は低い末梢抵抗に対して同じ駆出ボーラスを送る容積ポンプである。
左心室が広く研究されているように、右心室においても、収縮期血圧が著しく低く、駆出率がやや低い(55%)以外は、左心室の挙動と機能的に同一であると考えられてきた。
Revista Española de Cardiologíaの本号に掲載されたJuan C. Grignolaらの論文(37-42頁)1では、羊の心臓のソノミクロメータによる右心室、肺動脈、左心室、大動脈、肺流、心室径の圧力が研究されていて、この主張がいかに不正確であるかが示されています。 これらの著者によれば、右心室は駆出期が延長し、等容性弛緩期を欠くとのことである。
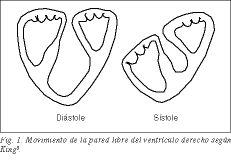
Torrentの概念1による肺動脈からの基部ループによって形成され、左心室の頂部ループを包含する右心室は、左心室の拡張であり、左心室の通常の機能に結合し条件づけられている。 三日月型の大きな表面積を持つため、低い肺抵抗に対して大量の血液を低コストで供給することができます。 犬で右心室の自由壁を切除しても、左心室の凸状中隔が右心室の駆出に積極的に協力するため、右心不全の兆候は現れないという古い経験も数多くある2-5。 右心室の自由壁を形成する帯が縦方向に短縮することで、収縮期に三尖弁が下がり、自由壁が蛇腹のように中隔の上で潰される。 1837年、ロンドンのガイズ病院の医師であったキングは、すでに中隔を固い壁と考え、自由壁は可動壁または延性壁と考えた (図1) 6 。 中隔は右腔側に膨らむことで、右心室の自由壁と協力して蛇腹効果を発揮する。
しかし右心室は、三尖弁から始まる流入部(洞)と肺動脈弁で終わる流出部(内腔)の2つの部分から構成されています。 Grignolaらの所見では、心室駆出は初期と後期の2段階で構成されていることが確認されている。 これは、解剖学的、発生学的、比較解剖学的、電気生理学的観察から、右心室の流入部と流出部は異なる部屋であり、さらに進化の異なる時期に出現したと考えられることと完全に一致するものであった。
内臓は心臓の流出路の筋肉化した部分で、系統的には古生代のシルル紀、4億3500万年前に顎のない環形動物(ヤツメウナギ)などの最も原始的な脊索動物に早くも現れている7 。 4億年前のデボン紀に、エラスモブランシュ亜綱のサメは、円形の繊維を持つ完全に筋肉化した内鰓孔(bulbus cordis)を持っている8 。 3億4500万年前の石炭紀ミシシッピア紀の現生両生類であるサンショウウオ(ウロデロ)では、肺内腔が非常に発達しており、肺狭窄の一部の例を彷彿させる。
一方、右心室の流入部はもっと新しく、古生代の石炭紀のペンシルバニア時代の爬虫類で、約2億7500万年前に現れ始め、初期の頂間中隔が出現し、おそらく空気を吸うときに全身循環と肺循環が分離することへの適応として現れた9 。 ワニでは、進化史上初めて、2つの静脈循環と動脈循環が無傷のまま完全に分離され、内膜は右心室に組み込まれている(鳥類や哺乳類では完全に組み込まれている)10 。 Torrent Guasp(私信)によれば、右心室の発達は、左心室の脱離と考えることができる(図2)。

Ontogeneticallyにも内果は非常に早い胚発生で、発生23-24日目の直心棒期に出現します。 管が湾曲し、右凸のループが形成されると、内槽を形成するbulbus cordisの尾部の拡張により右室洞が形成され、一方、心室間中隔は左右の2つの心室腔の拡張により受動的に「成長」する(27日目)11 。
また、いくつかの解剖学的観察から、右心室の洞と内腔は別個の構成要素であることが示されている。 右心室二室症は、右心室内流入路の狭窄が進行し、洞と内腔の間が閉塞する病態である。 単心室の最も一般的な形態であるダブルインレット左心室では、右室洞が存在せず、代わりに内腔が常に存在する。
電気生理学的研究でも、内膜の活性化は収縮期の比較的遅い時期に起こり、心臓のこの部分が最後に活性化することが示されており、その結果、右心室の流入・流出部分の収縮と緩和が非同期となる可能性がある12 。
Ginésらは羊で、収縮期において右心室では早期に最大駆出量に達し、駆出終了時間は左心室より有意に大きいことを示した。 つまり、右心室の収縮は、右心室の流入部から流出部(内槽)へと進む蠕動運動である。 この事実は、犬で示唆され、今回羊で実証されたものであるが、今回、人間でも証明されたのである。 音響定量心エコー、心音図、MRIの同時撮影により、右心室の流入部と内腔部には、流入部が収縮すると内腔部が拡大し、第二音の大動脈成分の後に収縮するような部位差があることが確認された13 。
この研究の第二の重要な事実は、右心室における等容性弛緩相の非存在を証明したことであり、私たちの考えでは、等容性収縮相もまた証明したことである。
拡張期に心室が充満するメカニズムについては、医学の歴史を通じて広く議論されてきた。 紀元前4世紀のErasistratusは、拡張期に心室が吸引ポンプとして機能すると考えていた。 しかし、ガレノス以来の収縮期と拡張期の概念を変えたハーヴェイは、『動物における心臓と血流の運動に関する解剖学的練習』の中で、「…血液が心室に入る方法は明白で、心臓(心室)が拡張して引き込むからではなく、耳介の脈(収縮)によってそこに放り込まれるから」14と述べている。 その後、Wiggersは、充血は主に拡張期の初めに起こることを考慮し、充血は等尺性弛緩期終了時の心房と心室の圧力差に依存するという結論に達した15。 しかし数年後、彼は「心室には弾性反動があり、心房収縮の前にほぼ完全に充満させることができる」と述べることができたのである16 。
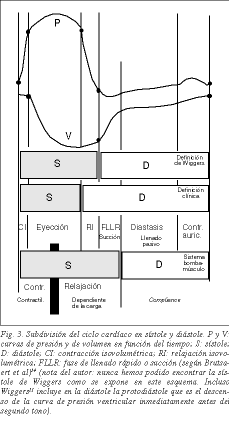
現代における活動的拡張期の概念は、Torrent Guaspの提案により、心室吸引の存在を証明することができたBrecherに始まる17 。 数年前、Dirk Brutsaertら18は、収縮期を現象論的ではなく概念的に理解し、等容性弛緩相と急速な心室充満相を伴う心弛緩を含むべきであると提案した(図3)。 実際、心臓が収縮前の状態に戻る過程と定義される弛緩は、能動的なプロセスである。 ATP消費は主に弛緩過程で二重経路で行われる。ミオシン頭部からのATPaseによるミオシンのアクチンからの剥離とSERCAによるトロポニンCからのCa+の小胞体への運搬、両方の過程が弛緩開始の合図となるのだ。 そのため、筋収縮のメカニズムにおける近年のマイルストーンの1つは、最大エネルギー消費の能動的なプロセスが弛緩であることを知ったことである19,20。 したがって、心周期はHarvey以来伝統的に行われてきたように収縮期と拡張期に分けることができるが、拡張期の最初の部分が活動的であること、すなわち等容弛緩期と急速な心室充満期または吸引期があるという知識は必要である。
吸引機構には多くの力が関わっており、右心室への銃創が左心房付属器を左心室内に侵襲するほど激しい吸引を起こした Roberts ら 21 の例を説明するには、その一部は非常に強力でなければならない。 心室らせんが弛緩する際の受動的な弾性反動が基本であると考えられるが、冠動脈が拡張期に充満する際の「血液供給の袖」の効果や、心室壁が拡張期に薄くなって半径が大きくなる際のラプラスの法則も作用しているものと思われる。 最近、Torrent Guaspは、心臓がピストンとして機能し、基部ループが頂点ループより伸縮することで排出が行われ(ピストンの下降)、吸引は基部ループを引きずる中隔の直線化による心室の伸長-拡張(ピストンの上昇)であると示唆した2 。
しかし、これは左心室で起こることです。 なぜ右心室では等容性弛緩が起こらず、したがって吸引も起こらないのでしょうか。 右心室の収縮があれば弛緩があり、駆出があれば充満があることは明らかです。 1つの可能性として、三角形の右室容積-圧力曲線は、内腔の収縮とはるかに大きな流入部の充満という、2つの異なる事象を同時に記録していることが考えられる。 この考えを支持する論文は少なくとも2つある。 March ら 21 は右室が独立した 2 室からなることを示唆し、Pouleur ら 22 は自由壁が弛緩しているときにも右室駆 出が継続することを確認した。これは聴診の観点から Shaver らによって既に提唱されており、2 音の肺成分の遅れを説明するために「絞扼」と名づけられたものである。
もう一つの可能性は、肺動脈が低インピーダンスであるため、吸引段階を必要とせず、単純な房室圧低下で右心充満を行うというものである。 この単純な考え方に賛成なのは、もし等容性弛緩相がなければ等容性収縮相も必要ないということである。
吸引を起こすには等容性収縮と等容性弛緩の相が必要なのは、高残荷のせいでしょうか。
いずれにせよ、右室容積-圧力曲線の三角形と左室容積-圧力曲線の二次曲線との違いは、魅力的で客観的な事実であり、その血行動態的解釈はさらなる考察と研究が必要である。