4 wesentliche Nebenwirtel einer Blume | Botanik
Hinweise:
Die folgenden Punkte heben die vier wesentlichen Nebenwirtel einer Blume hervor. Die wesentlichen Nebenwirtel sind:- 1. Kelch 2. Corolla 3. Androeceum 4. Gynoeceum.
Zusatzwirtel # 1. Kelch:
Der Kelch ist der äußerste akzessorische Satz von Blütenblättern und besteht aus Kelchblättern. Kelchblätter sind meist grüne, unscheinbare Körper. Sie schützen die wesentlichen Quirle im Knospenstadium vor mechanischen Verletzungen und Austrocknung.
Sie haben also hauptsächlich eine Schutzfunktion. Bei einigen Pflanzen sind die Kelchblätter wie die Blütenblätter gefärbt und werden petaloid genannt, z. B. bei Caesalpinea (B. Krishnachura). Bei Mussaenda ist eines der Kelchblätter in eine flache, blattähnliche Struktur umgewandelt (Abb. 88). Bei den Sonnenblumengewächsen sind die Kelchblätter zu haarigen Körpern, den Pappus, umgebildet, die bei der Verbreitung der Früchte helfen.
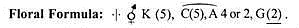
Wenn die Kelchblätter frei sind, nennt man den Kelch polysepal, sind sie vereinigt, ist der Kelch gamosepal. Der gamosepalige Kelch gleicht mehr oder weniger einer Röhre, in der die Anzahl der Kelchblätter durch Zählen der Zähne oder Lappen leicht bestimmt werden kann. In vielen Fällen fallen die Kelchblätter ab, bevor sich die Blüten vollständig öffnen. Man nennt sie kaduzierte Kelchblätter, z. B. beim Mohn, während sie bei anderen Pflanzen nach der Befruchtung mit anderen Teilen abfallen.
Dies sind sommergrüne Kelchblätter. Bei manchen Blüten bleiben die Kelchblätter sogar bis zur Frucht bestehen und werden als persistent bezeichnet, z. B. bei Brinjal, Tomate. Manchmal bleiben sie nicht nur bestehen, sondern wachsen zu einem Teil der Frucht, wie bei Dillenia (B. Chalte), Physalis (B. Tepari). Diese werden als heranwachsende Kelchblätter bezeichnet.
Zusatzquirle Nr. 2. Corolla:
Sie ist der zweite Nebenquirl der Blüte, dessen einzelne Teile als Blütenblätter bezeichnet werden. Sie machen die Blüten auffällig und attraktiv, denn die Blütenblätter sind leuchtend gefärbt und duften oft süßlich.
Die Funktion der Blumenkrone ist zweifach:
SCHUTZ:
Erstens, der Schutz der essentiellen Quirle im Knospenstadium; und
Zweitens, das Anlocken von Insekten zur Bestäubung.
Die Blütenblätter von Polyalthia (B. Debdaru), Puddingapfel usw. werden sepaloid genannt, da sie grün und sepalartig sind. Sind die Blütenblätter frei, so nennt man die Blumenkrone polypetal; sind sie vereinigt, so ist sie gamopetal.
Die freien Blütenblätter bestehen oft aus zwei Teilen – einem schmalen Stiel, der als Kralle bezeichnet wird, und einem oberen flachen Blatt, das als Glied bezeichnet wird. Blütenblätter, die Krallen haben, werden als krallenförmig bezeichnet, wie bei Rosa, Rettich, Senf (Abb. 89, 1). Die Blumenkrone ist regelmäßig, wenn alle Blütenblätter in Größe und Form ähnlich sind; sind sie unähnlich, ist sie unregelmäßig.
Formen der Blumenkrone:
Es gibt verschiedene Formen von Blumenkronen in Blumen. Sie können frei oder vereinigt, regelmäßig oder unregelmäßig sein. Es ist keineswegs leicht, alle in der Natur vorkommenden Formen genau zu beschreiben.
Die folgenden sind die wichtigsten und häufigsten Formen (Abb. 89):
A. Polypetalous Regular:
1. Kreuzförmig:
Sie besteht aus vier freien, krallenförmigen Blütenblättern, die in Form eines Kreuzes angeordnet sind (Abb. 89, 1). Beispiele – Senf, Rettich. Sie ist charakteristisch für die Familie der Senfgewächse (Cruciferae).
Hinweise:
2. Garyophyllaceous:
Sie hat fünf krallenförmige Blütenblätter, deren Glieder mehr oder weniger rechtwinklig zu den Krallen liegen, z.B. rosa (Abb. 89, 2).
Hinweise:
3. Rosafarben:
Sie besteht aus fünf Blütenblättern ohne Krallen, z. B. Rosen, Tee (Abb. 89, 3).
B. Polypetalous Unregelmäßig:
1. Papilionaceous:
Hinweise:
Sie sieht mehr oder weniger aus wie ein Schmetterling (papilion=Schmetterling). Sie besteht aus fünf freien Blütenblättern. Das größte und äußerste wird Fahne oder Vexillum genannt; zwei seitliche heißen Flügel oder Alae; und die beiden innersten, Kiel oder Carina genannt, vereinigen sich zu einem bootsförmigen Körper. Beispiele – Erbse (Abb. 89, 4 & 5), Bohnen, Sesbania (B. Bak). Es ist charakteristisch für die Familie der Erbsengewächse (Papilionaceae).
C. Gamopetalous Regular:
1. Röhrenförmig:
Hier vereinigen sich die Blütenblätter zu einem röhrenförmigen Körper mit Lappen oder Zähnen an der Spitze, durch deren Auszählung die Anzahl der Blütenblätter bestimmt wird, z. B. Scheibenblüten der Sonnenblume, Ringelblume (Abb. 89, 6).
Abkürzungen:
2. kelchförmig:
Fünf Blütenblätter vereinigen sich zu einer glockenförmigen Blumenkrone, wie beim Kürbis (Abb. 89, 7), Physalis (B. Tepari).
3. Infundibuliform:
Sie besteht aus fünf Blütenblättern, die sich zu einer trichterförmigen, sich zur Spitze hin allmählich erweiternden Blumenkrone vereinigen, z. B. Datura (Abb. 89, 8), Thivetia (B. Kalke).
4. Drehen:
Hier ist die Röhre kurz und die spreizenden Glieder sind mehr oder weniger rechtwinklig zur Röhre angeordnet, wodurch die Blumenkrone radförmig erscheint (Abb. 89, 9), z. B. Brinjal, Nyctanthes (B. Shieuli).
Hinweise:
5. Hypokraterförmig:
Sie ist wie die drehbare, hat aber eine vergleichsweise lange Röhre (Abb. 89, 10), z. B. Vinca (B. Nayantara).
D. Gamopetalous Unregelmäßig:
1. Bilabiate oder Zweilippige:
Hier bilden die Blütenblätter durch Vereinigung zwei Teile mit einem offenen weiten Mund zwischen ihnen. Der untere Teil besteht aus drei, der obere aus zwei Blütenblättern (Fig. 89, 11), z. B. Ocimum (B. Tulsi), Leonurus (B. Drone).
2. Personate:
Abweichungen:
Es ist wie das bilabiate, aber der Mund ist geschlossen, da die beiden Lippen sehr nahe beieinander liegen, z. B. Löwenmäulchen (Abb. 89, 12), Lindenbergia (B. Basanti).

3. ligulär oder riemenförmig:
Die fünf Blütenblätter sind am Grunde zu einer kurzen Röhre vereinigt, breiten sich aber oben wie ein Band aus, z. B. Zungenblüten der Sonnenblume, Ringelblume.

Anhängsel der Blumenkrone:
Die Blumenkrone ist oft mit Spornen oder langen Fortsätzen versehen, wie beim Springkraut (B. Dopati), und mit Auswüchsen, die Corona genannt werden und die Schönheit der Blüten erhöhen. Die Corona ist bei der Passionsblume (B. Jhumko) als schöner Quirl von Auswüchsen zwischen der Corolla und dem Androeceum vorhanden.
Nektarien, die honigabsondernden Drüsen, sind manchmal an der Basis der Blütenblätter vorhanden. Sie sind natürliche Hilfsmittel, um Insekten zur Bestäubung anzulocken.
Aestivation:
Aestivation ist die Art der Anordnung von Kelch- und Kronblättern im Knospenstadium zueinander (Abb. 90).

Es gibt die folgenden Typen:
(i) Valvat, wenn sich die Glieder entweder berühren oder sehr eng nebeneinander liegen, aber nicht überlappen, wie bei Calotropis (B. Akanda), Artabotrys (B. Kantali champa).
(ii) Gedreht oder verdreht, wenn die Glieder so angeordnet sind, dass sich jedes von ihnen an einem Rand überlappt und das nächste Glied am anderen Rand überlappt, wie bei der China-Rose.
(iii) Schuppenförmig. Es handelt sich um eine Abwandlung der verdrehten Anordnung, bei der ein Glied ganz außen ist, d.h. von beiden Rändern überlappt wird, ein Glied ganz innen ist, d.h. von beiden Rändern überlappt wird und der Rest wie bei der verdrehten Ästivation angeordnet bleibt, z.B. Cassia (B. Kalkasunde).
(iv) Vexillar: Hier ist von den fünf Blütenblättern das größte das äußerste. Es schließt zwei Seitenblätter ein, die ihrerseits die beiden innersten umschließen. Sie ist charakteristisch für Schmetterlingsblütler wie Erbse, Sesbania (B. Bak).
Perianth:
Die akzessorischen Quirle von Kelch und Blumenkrone werden oft als Perianth bezeichnet. Bei den Einkeimblättrigen ist das Perianth nicht in zwei Gruppen unterteilt, wie bei der Schlauchrose, Crinum (B. Sukhadarsan) usw. Wenn die Lappen frei sind, ist das Perianth mehrblättrig, wenn sie vereinigt sind, ist es gammophyll. Bei den Gräsern wird das Perianth durch ein Paar kleiner Schuppen dargestellt, die Lodicula genannt werden.
Zusatzquirl Nr. 3. Androeceum:
Das Androeceum ist der dritte Satz und der erste wesentliche Wirtel der Blüte. Es besteht aus Staubblättern oder Mikrosporophyllen, die männliche Organe sind. Ein Staubblatt besteht normalerweise aus zwei Teilen, nämlich einem schlanken Stiel, der Staubfaden genannt wird, und einem geschwollenen Körper an der Spitze des Staubfadens, der als Anthere bezeichnet wird.
Die Anthere hat gewöhnlich zwei Lappen an den beiden Seiten; jeder Lappen wiederum hat zwei sackartige Körper, die Pollensäcke oder Mikrosporangien genannt werden und unzählige kleine einzellige Pollenkörner oder Mikrosporen enthalten.
Bei der China-Rose ist in jedem Staubbeutel ein Pollensack vorhanden. Die Verlängerung des Fadens, der die beiden Staubbeutel zusammenhält oder verbindet, wird Konnektiv genannt. Durch das Konnektivum verläuft ein Gefäßbündel, das die Nahrung zur wachsenden Anthere transportiert.
Nicht alle Staubblätter tragen immer fruchtbare Antheren. Bei Cassia sophera (B. Kalkasunde) sind vier von zehn Staubblättern steril. Sie werden Staminodien genannt. Bei Canna (B. Sarbajaya) sind die Staminodien gefärbt und sehen blütenblattartig aus. Bei Calotropis (B. Akanda) finden sich Auswüchse an den Staubblättern, die als Staubblattkranz bezeichnet werden.
Befestigung der Staubfäden an den Staubbeuteln:
Die Staubgefäße werden nach der Art der Befestigung der Staubbeutel an den Staubfäden in folgende Typen eingeteilt:
(i) Basisfixiert oder angeboren, wenn der Staubfaden nur an der Basis der Antheren befestigt ist, wie beim Senf (Abb. 91).

(ii) Adnat, wenn aufgrund des Vorhandenseins eines gut ausgeprägten Konnektivs der Faden über die gesamte Länge der Antheren zu verlaufen scheint, z. B. bei Michelia (B. Champaka).
(iii) Dorsifixiert, wenn der Faden an der Rückseite der Anthere befestigt ist, wie bei der Passionsblume, Sesbania (B. Bak).
(iv) Vielgestaltig, wenn der Faden an der Rückseite der Anthere durch eine scharfe Spitze befestigt ist, so dass die Anthere frei in der Luft schwingen kann, z. B. Gräser, Paddy.
Dehiszenz der Antheren:
Bei der Reife der Antheren verschmelzen die beiden Pollensäcke, da die Wand zwischen ihnen aufbricht. Die Pollenkörner verbleiben dann verstreut im Hohlraum.
Die Antheren dehiszieren oder platzen, um die Pollenkörner freizugeben, so dass sie durch verschiedene Agentien zu den Narben der Blüten getragen werden können. Die Dehiszenz erfolgt meist durch Längsspaltung, wie beim Stechapfel, durch Poren, die sich an den Spitzen der Antheren bilden, wie bei der Brinjal, oder durch eine Reihe von Klappen (Valven), wie beim Zimt.
Relative Länge der Staubblätter:
Die Staubblätter einer Blüte können gleich oder ungleich lang sein. Ipomoea (B. Kalmi) hat fünf Staubblätter, die alle ungleich lang sind. Die Blüten der Senfgewächse haben gewöhnlich sechs Staubblätter in zwei Quirlen.
Zwei der äußeren Quirle sind kurz und vier der inneren Quirle haben lange Staubfäden. Sie werden als tetradynamische Staubblätter bezeichnet (Abb. 91). Leonurus (B. Drone), Ocimum (B. Tulsi), etc. haben vier Staubblätter, von denen zwei lang und zwei kurz sind. Sie werden didynamische Staubblätter genannt.
Vereinigung der Staubblätter:
Die Staubblätter können sich untereinander oder mit anderen Quirlen vereinigen. Die Vereinigung gleichartiger Glieder nennt man Kohäsion, diejenige ungleichartiger Glieder wird als Adhäsion bezeichnet.
Kohäsion der Staubblätter:
Da die Staubblätter aus zwei Hauptteilen bestehen – Filamenten und Staubbeuteln -, sind zwei Zustände des Zusammenhalts möglich:
(i) Die Staubblätter können sich durch die Filamente vereinigen, die Antheren bleiben frei; oder
(ii) sie können durch die Antheren vereinigt werden, die Filamente bleiben frei (Abb. 92).
Die Vereinigung durch Staubfäden führt zur Bildung eines Bündels oder einer Adelphie. Wird ein solches Bündel durch Vereinigung von Staubfäden gebildet, so nennt man die Staubblätter einblütig, wie bei der Chinarose, der Baumwolle.
Wenn zwei Bündel gebildet werden, sind sie diadelphisch, wie bei Erbse, Sesbania (B. Bak); und wenn mehr als zwei Bündel gebildet werden, nennt man sie polyadelphisch, wie bei Seide-Baumwolle. Wenn sie sich dagegen an den Staubbeuteln vereinigen, wobei die Staubfäden frei bleiben, nennt man sie syngenesisch, wie bei Kürbis, Sonnenblume.

Anhaftung der Staubblätter:
Die Staubblätter können sich mit dem nächstunteren Wirtel, d.h. der Blumenkrone oder dem Perianth, oder mit dem nächstoberen Wirtel, dem Stempel, vereinigen. Wenn sie sich mit den Blütenblättern vereinigen, nennt man die Staubblätter epipetal.
Fast alle Blüten mit gamopetaler Blumenkrone haben epipetale Staubblätter. Wenn sie an den Perianthlappen anhaften, nennt man die Staubblätter epiphyllisch, wie bei der Röhrenrose. Die Staubblätter werden gynandrisch genannt, wenn sie am Griffel anhaften, z. B. bei Calotropis (B. Akanda).
Pollenkörner:
Die Pollensäcke oder Mikrosporangien der Antheren enthalten einige spezielle Zellen, die Pollen-Mutterzellen oder Mikrosporen-Mutterzellen genannt werden. Jede von ihnen macht eine Reduktionsteilung durch, um eine Tetrade von Pollenkörnern oder Mikrosporen zu erzeugen. Diese sind kleine, staubige, einzellige Körper von meist runder oder ovaler Form. Ein Pollenkorn hat Protoplasma mit einem einzigen Kern, der von zwei Hüllen umgeben ist.
Die innere Hülle, Intine genannt, ist dünn, zart und besteht aus Zellulose; die äußere Hülle, Exine genannt, ist dick, kutinisiert und oft mit warzigen oder stacheligen Fortsätzen versehen. Auf der Exine befinden sich hier und da einige unverdickte Stellen oder Schwachstellen, die Keimporen genannt werden.
Die unbefruchteten Pollenkörner werden im Allgemeinen zweikernig, bevor sie sich aus den Beuteln lösen. In einigen Blüten sind die Pollenkörner, statt lose zu bleiben, zu Massen, die Pollinien genannt werden, zusammengeballt, wie bei Calotropis (B. Akanda), Orchideen.
Zubehör Quirle # 4. Gynoeceum oder Griffel:
Das Gynoeceum oder der Griffel ist der letzte und innerste Satz von Blütenblättern, der die zentrale Position auf dem Thalamus einnimmt. Es besteht aus Karpellen oder Megasporophyllen, die die weiblichen Organe der Blüten sind.
Ein Karpell hat gewöhnlich drei Teile:
(i) Der basale geschwollene Teil ist der Fruchtknoten,
(ii) der schlanke zylindrische Bereich ist der Griffel, der gewöhnlich in der rezeptiven Stelle endet
(iii) Narbe.
Der Fruchtknoten enthält einen oder mehrere eiförmige Körper, die Eizellen genannt werden und nichts anderes als unreife Samen sind. Der Griffel kann aus einem, zwei, drei oder vielen Fruchtblättern bestehen. Dementsprechend sind sie monokarpellar, bikarpellar, trikarpellar oder polykarpellar. Der monokarpellare Griffel wird als einfach bezeichnet, während die anderen als zusammengesetzt bezeichnet werden.
Die blattartige Natur eines Fruchtknotens zeigt sich in einem einfachen Griffel, der mit einem einfachen Blatt verglichen werden kann. Wenn das Blatt entlang der Mittelrippe gefaltet wird, treffen die beiden Ränder aufeinander und bilden einen Hohlraum, der einem einfachen Griffel ähnelt.
Die Linie, an der die beiden Ränder zusammentreffen, wird als Ventralnaht bezeichnet, diejenige, die der Mittelrippe entspricht, ist die Dorsalnaht. Entlang der Ventralnaht bilden sich weiche kissenartige Körper oder Leisten, die Plazenta genannt werden und an denen die Samenanlagen haften bleiben.
Der zusammengesetzte Griffel wird apokarp genannt, wenn die Fruchtblätter frei sind. Hier bleiben alle Fruchtblätter auf dem Thalamus sitzen, jedes hat seinen eigenen Fruchtknoten, Griffel und Narbe, wie bei Michelia (B. Champa), Artabotrys (B. Kantali champa).
Sind aber die Fruchtblätter vereinigt, so nennt man den Stempel synkarpatisch, wo gewöhnlich ein gemeinsamer Fruchtknoten und Griffel gebildet wird, wobei die Fruchtblätter gewöhnlich nur in der Gegend der Narben frei bleiben, z. B. China-Rose (Fig. 98), Baumwolle, Datura.

Der Griffel ist normalerweise die schlanke zylindrische Fortsetzung des Fruchtknotens. Bei der Familie Ocimum (B. Tulsi) ist der Fruchtknoten vierlappig und der Griffel entspringt an der Basis des Fruchtknotens oder am Thalamus. Diese Art von Griffel wird als gynobasisch bezeichnet (Abb. 93). Gewöhnlich sind die Griffel laublos, aber bei Mais finden sich auch seidige, ausdauernde Griffel.

Die Narbe ist die endständige rezeptive Stelle. Sie kann flach und klebrig sein, weil sie Narbenflüssigkeit absondert, und bei einkeimblättrigen Pflanzen federartig. Dies sind Vorrichtungen, die den Empfang der Pollenkörner erleichtern.
Plazentation:
Der Fruchtknoten kann unilokular [uni = eine, loculus = Kammer) oder multilokular (multi = viele, loculus = Kammer) sein. Die Eizellen oder Mega-Sporangien bleiben an den polsterartigen parenchymatösen Ausstülpungen des Fruchtknotens, der Plazenta, haften, und die Art der Anordnung der Plazenta wird als Plazentation bezeichnet.
Typen der Plazentation (Abb. 94):
1. Marginal:
Die marginale Plazentation ist charakteristisch für die Familie der Erbsengewächse, bei denen der Griffel einfach ist und die Plazenta mit den Eizellen an der Bauchnaht liegt.
2. Parietal:
Hier ist der Griffel mehrkarpellig und die Plazenta liegt in den Bereichen, wo sich die Ränder der Karpellen treffen. Der Fruchtknoten ist unilokulär und die Plazentagruppen entsprechen der Anzahl der vorhandenen Fruchtblätter, z.B. Papaya, Kürbis.
3. Axil:
Bei der Axil-Plazentation ist auch der Griffel polykarpellar. Die Ränder der Fruchtblätter laufen nicht zusammen, sondern biegen sich nach unten und bilden in der Mitte eine Achse. Die Plazenta, die die Samenanlagen trägt, ist um die Achse herum verteilt. So wird der Fruchtknoten vielsamig, wobei die Zahl der Fruchtknoten gleich der Zahl der vorhandenen Fruchtblätter ist, z. B. Orange, China-Rose, Stechapfel.
4. Free Central:
Hier sind die Plazenta um die Achse herum vorhanden, wie bei der Axillenplazentation, aber durch die Absorption der Scheidewände wird der multilokulare Fruchtknoten unilokular. Manchmal sind Reste der Scheidewände zu sehen. Beispiel-rosa
5. Oberflächlich:
Es handelt sich ebenfalls um einen vielkammerigen Fruchtknoten, aber die Plazenta mit den Eizellen sind überall auf der Innenfläche der Trennwände vorhanden, wie bei der Seerose.
6. Basal:
Hier ist der Fruchtknoten einhäusig, und die Plazenta mit einer einzelnen Samenanlage entwickelt sich direkt am Thalamus an der Basis des Fruchtknotens, z. B. Sonnenblume, Dahlie.

Die Eizelle:
Die Samenanlage oder Mega-Sporangium entwickelt sich nach der Befruchtung zum Samen. Jede Eizelle ist ein eiförmiger Körper mit einem Stiel, der Funiculus genannt wird, mit dem sie an der Plazenta befestigt bleibt.
Der weiche Hauptkörper der Eizelle wird als Nucellus bezeichnet, der von zwei Hüllen, den Integumenten, umgeben bleibt. Die äußeren und inneren Integumente umschließen die Eizelle fast vollständig und lassen nur eine kleine Öffnung, die Mikropyle, an der Spitze übrig. Bei manchen Eizellen ist nur ein Integument vorhanden.
Die breite Basalregion, wo die Integumente auf den Nucellus treffen, wird als Chalaza bezeichnet. Ein ovaler Körper, der gescheiterte Embryosack, ist zum mikropylaren Ende hin in den Nucellus eingebettet. Der Embryosack hat normalerweise acht Kerne. Drei von ihnen sammeln sich am mikropylaren Ende und vereiteln den Eiapparat.
Der mittlere der drei Kerne ist das Ei oder die weibliche Gamete, die beiden anderen werden Hilfszellen oder Synergiden genannt. Drei, die sich am chalazalen Ende sammeln, werden als antipodale Zellen bezeichnet; und die beiden übrigen, die polaren Kerne genannt werden, liegen entweder nebeneinander oder verschmelzen in der zentralen Region und bilden den so genannten definitiven Kern (Abb. 95). Der Embryosack ist der wichtigste Teil der Eizelle.

Bildung der Eizelle und des Embryosacks:
Die Eizelle entsteht als kleine rundliche Ausstülpung von Weichgewebe (Nucellus) auf der Plazenta. Allmählich entwickeln sich kragenartige Auswüchse oder Integumente, die den Nucellus bedecken und an der Spitze eine Öffnung, die Mikropyle, lassen.
Vor der vollständigen Ausbildung der Integumente wird eine hypodermale Zelle an der Spitze des Nucellus auffallend. Dies ist die Megasporen-Mutterzelle. Sie unterliegt einer Reduktionsteilung, aus der vier Zellen in einer Reihe entstehen, was als lineare Tetrade bezeichnet wird.
Drei von ihnen zerfallen im Allgemeinen, und die innerste überlebt. Der Kern der funktionsfähigen Megaspore teilt sich nun durch Mitose in zwei, die zu den beiden Enden der Megaspore wandern. Jeder von ihnen teilt sich wiederum zweimal. So dass an jedem Ende vier Kerne vorhanden sind.
Drei von ihnen am mikropylaren Ende bilden den Eiapparat mit dem zentralen hervorstehenden Ei und den beiden Synergiden. Drei, die am chalazalen Ende aggregiert sind, bilden die antipodalen Zellen. Die verbleibenden zwei, oft als polare Kerne bezeichnet, wandern in die Mitte der Megaspore und bilden den endgültigen Kern (Abb. 96).

Dies ist ein Abriss des Prozesses der normalen Art der Entwicklung des Embryosacks bei den Angiospermen.
Formen der Eizellen:
Die Eizellen sind von folgenden Typen (Abb. 97):
(1) Orthotrop oder gerade, wenn die Samenanlage aufrecht ist und der Funiculus, die Mikropyle und die Chalaza in einer geraden Linie liegen. Dabei liegt die Mikropyle zum Apex und die Chalaza zur Basis hin.
(2) Anatrop oder umgedreht, wenn die Samenanlage durch Überwachsen des Funiculus umgedreht ist, so dass die Mikropyle an der Basis in der Nähe des Funiculus und die Chalaza am anderen Ende liegt. Dieser Typ ist am häufigsten.
(3) Amphitrop oder quer, wobei der Hauptkörper der Samenanlage mehr oder weniger im rechten Winkel zum Funiculus steht, z. B. Lemna (B. Kshudi-pana).
(4) Campylotrop oder gekrümmt, wobei der Hauptkörper der Samenanlage gekrümmt oder hufeisenförmig gebogen ist und mehr oder weniger rechtwinklig zum Funiculus angeordnet ist, z. B. Senf.

Blütendiagramm:
Die in einer Blüte vorhandenen Teile und ihre Anordnung zueinander können durch Diagramme dargestellt werden, die Blumendiagramme genannt werden. Sie bestehen aus einer Anzahl konzentrischer Ringe, die den vorhandenen Blütensätzen entsprechen und in denen vereinigte und freie Teile dargestellt werden können. Die Position der Blütenachse wird durch einen „Punkt“ dargestellt. Die beste Vorstellung von einem Blütendiagramm erhält man, wenn man eine Blütenknospe quer durchschneidet.
Blütenformeln:
Blütenformeln stellen die Teile einer Blüte mit ihrer jeweiligen Position dar. Dabei werden einige Zeichen und Symbole verwendet. K steht für Kelch, C für Blumenkrone, P für Perianth, A für Androeceum und G für Gynoeceum. Die Vereinigung wird durch das Setzen der Buchstaben in Klammern dargestellt, die Verwachsung durch Linien über den betreffenden Wirteln.
Mitglieder desselben Satzes, die in zwei Wirteln vorhanden sind, werden durch das Zeichen „+“ (plus) zwischen ihnen dargestellt, co steht für viele Mitglieder in einem Wirtel. Der obere oder untere Fruchtknoten wird durch horizontale Linien unter oder über der Zahl der Fruchtblätter angegeben.
Nachfolgend sind die Formeln einiger häufiger Blüten aufgeführt: 