Hemodynamische verschillen tussen de rechter- en de linkerhartkamer | Revista Española de Cardiología
De linkerhartkamer is een drukpomp die een uitwerpbolus stuurt tegen hoge perifere weerstanden, terwijl de rechterhartkamer een volumepomp is die dezelfde uitwerpbolus stuurt tegen lage perifere weerstanden.
Zoals de linker ventrikel uitvoerig is bestudeerd, is er bij de rechter ventrikel van uitgegaan dat deze, afgezien van de duidelijk lagere systolische druk en de iets lagere ejectiefractie (55%), qua functie identiek is aan het gedrag van de linker ventrikel.
Het in dit nummer van de Revista Española de Cardiología gepubliceerde artikel van Juan C. Grignola e.a. (blz. 37-42)1 , waarin de drukken van de rechterhartkamer, de longslagader, de linkerhartkamer, de aorta, de longstroom en de ventriculaire diameters door middel van sonomicrometrie in schapenharten worden bestudeerd, toont aan hoe onnauwkeurig deze bewering is. Volgens deze auteurs heeft de rechterventrikel een verlengde ejectiefase en ontbreekt de isovolumische relaxatiefase.
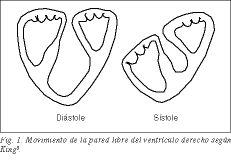
De rechterkamer, gevormd door de basale lus van de longslagader, volgens het concept van Torrent 1 , die de apicale lus van de linkerkamer omsluit, is een uitbreiding van de linkerkamer, gekoppeld aan en geconditioneerd door de normale werking van de linkerkamer. Door zijn grote halvemaanvormige oppervlak kan hij grote hoeveelheden bloed afgeven tegen lage longweerstanden en tegen lage kosten. Er zijn talrijke oude ervaringen dat, door het verwijderen van de vrije wand van de rechterhartkamer bij de hond, er geen tekenen van rechterhartfalen optreden, omdat het bolle septum van de linkerhartkamer actief meewerkt aan de uitdrijving van de rechterhartkamer 2-5 . De verkorting in de lengterichting van de band die de vrije wand van de rechterventrikel vormt, resulteert in systolische verlaging van de tricuspidalis annulus en verplettering van de vrije wand over het septum als een blaasbalg. Reeds King, een arts in het Guy’s Hospital in Londen in 1837, beschouwde het septum als een massieve wand, terwijl de vrije wand een beweeglijke of buigzame wand zou zijn (fig. 1) 6 . Het septum, dat naar de rechterholte toe uitpuilt, werkt samen met de vrije wand van de rechterhartkamer in het balgeffect.
Maar de rechterhartkamer bestaat uit twee delen: een instroomgedeelte of sinus dat begint bij de tricuspidalisklep en een uitstroomgedeelte of infundibulum dat eindigt bij de pulmonaalklep. In de bevindingen van Grignola et al. wordt geconstateerd dat de ventriculaire ejectie uit twee fasen bestaat, een vroege en een late. Dit is volledig in overeenstemming met anatomische, embryologische, vergelijkende anatomische en elektrofysiologische waarnemingen, die erop wijzen dat het instroom- en het uitstroomgedeelte van de rechterhartkamer verschillende kamers zijn, die bovendien in verschillende perioden van de evolutie zijn verschenen.
Het infundibulum is een gespierd deel van het uitstroomtraject van het hart dat fylogenetisch al verschijnt bij de meest primitieve chordaten zoals de kaakloze cyclostomen (lamprei) in het Siluur van het Paleozoïcum, 435 miljoen jaar geleden 7 . In het Devoon, 400 miljoen jaar geleden, hebben haaien van de subklasse elasmobranchii een volledig gespierd infundibulum (bulbus cordis) met cirkelvormige vezels 8 . Bij de salamander (urodelo), een modern amfibie uit het Mississippien van het Carboon 345 miljoen jaar geleden, is het infundibulum zo ontwikkeld dat het doet denken aan sommige gevallen van pulmonale stenose.
Het instroomgedeelte van de rechter hartkamer daarentegen is veel recenter, en begint te verschijnen bij reptielen uit het tijdperk Pennsylvanië van het Carboon, ook van het Paleozoïcum, ongeveer 275 miljoen jaar geleden, wanneer het beginnende apicale interventriculaire septum ontstaat, mogelijk als een aanpassing aan de scheiding van de systemische en pulmonale circulatie bij het inademen van lucht 9 . Bij de krokodil zijn voor het eerst in de evolutionaire geschiedenis de twee veneuze en arteriële circulaties nu volledig gescheiden met een intact interventriculair septum, en is het infundibulum opgenomen in de rechter ventrikel, hoewel volledige opname voorkomt bij vogels en zoogdieren 10 . De ontwikkeling van de rechterkamer kan worden beschouwd als een dehiscentie van de linkerkamer, volgens Torrent Guasp (persoonlijke mededeling) (fig. 2).

Ontogenetisch gezien is het infundibulum ook van zeer vroege embryologische ontwikkeling, en verschijnt het in het rechte hartbuisstadium op 23-24 dagen van ontwikkeling. Wanneer de buis incurved is en de rechter convexe lus is gevormd, wordt de rechter ventrikel sinus gevormd door expansie van het caudale deel van de bulbus cordis, die het infundibulum vormt, terwijl het interventriculaire septum passief “groeit” door expansie van de twee ventriculaire holten aan weerszijden (dag 27) 11 .
Verscheidene anatomische observaties tonen eveneens aan dat de sinus en het infundibulum van de rechter ventrikel afzonderlijke componenten zijn. De dubbelkamer rechterventrikel is een laesie die wordt gekenmerkt door een progressieve stenose van de instroom van het infundibulum, die leidt tot obstructie tussen de sinus en het infundibulum. Bij dubbelinlaat linkerventrikel, de meest voorkomende vorm van enkelvoudige ventrikel, is de rechter ventrikel sinus afwezig en is in plaats daarvan altijd het infundibulum aanwezig.
Elektrofysiologische studies wijzen er ook op dat activering van het infundibulum relatief laat in de systole optreedt en dat dit deel van het hart als laatste wordt geactiveerd, wat kan resulteren in asynchrone contractie en relaxatie van het instroom- en uitstroomgedeelte van de rechter ventrikel 12 .
Ginés et al demonstreren bij schapen dat in de systole, in de rechter ventrikel, de maximale ejectie vroeg wordt bereikt en dat het einde van de ejectie beduidend groter is dan in de linker ventrikel. Met andere woorden, de samentrekking van de rechterhartkamer is een peristaltische samentrekking die verloopt van het instroom- naar het uitstroomgedeelte (infundibulum) van de rechterhartkamer. Dit feit, dat bij de hond werd gesuggereerd en in dit werk bij schapen wordt aangetoond, is nu ook bij de mens bewezen. Door middel van gelijktijdige akoestische gekwantificeerde echocardiografie, fonocardiografie en magnetische resonantie beeldvorming is bevestigd dat er regionale verschillen zijn tussen het instroomgedeelte en het infundibulum van de rechter hartkamer, zodanig dat het infundibulum uitzet wanneer het instroomgedeelte samentrekt en samentrekt na de aortacomponent van de tweede toon 13 .
Het tweede belangrijke feit van dit werk is de demonstratie van het niet-bestaan van de isovolumetrische relaxatiefase in de rechterventrikel, en naar onze mening, ook van de isovolumetrische contractiefase.
Het mechanisme waarmee de ventrikels zich vullen in diastole is veelbesproken in de geschiedenis van de geneeskunde. Erasistratus in de 4e eeuw v. Chr. geloofde dat het ventrikel tijdens de diastole als een zuigpomp fungeerde. Harvey echter, die de sinds Galen gangbare begrippen systole en diastole veranderde, zei in zijn “Exercitatio anatomica de motu cordis et sanguinis in animalibus”: “…de manier waarop het bloed in de hartkamers komt is duidelijk: niet omdat het hart (ventrikel) het naar binnen trekt omdat het verwijdt, maar omdat het erin wordt geworpen door de polsslag (contractie) van de oorschelpen” 14 . Later kwam Wiggers, rekening houdend met het feit dat vulling voornamelijk optreedt aan het begin van de diastole, tot de conclusie dat vulling afhing van de drukverschillen tussen boezem en ventrikel aan het eind van de isometrische relaxatiefase 15 . Jaren later kon hij echter verklaren dat “de hartkamers een elastische terugslag hebben die hen in staat stelt zich bijna volledig te vullen vóór de atriale contractie” 16 .
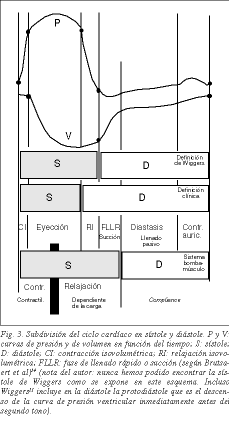
Het moderne concept van actieve diastole begint met Brecher die, op suggestie van Torrent Guasp, het bestaan van ventriculaire suctie kon aantonen17 . Enkele jaren geleden stelden Dirk Brutsaert et al18 voor de systole vanuit een conceptueel en niet vanuit een fenomenologisch oogpunt te begrijpen en deze de cardiale relaxatie met isovolumetrische relaxatiefasen en de snelle ventriculaire vulfase te laten omvatten (fig. 3). Ontspanning, gedefinieerd als het proces waarin het hart terugkeert naar de precontractiele configuratie, is namelijk een actief proces. ATP-verbruik wordt voornamelijk gebruikt in het relaxatieproces via een tweeledige route: ATP-verbruik door ATPase van myosinekoppen om myosine los te maken van actine en door SERCA’s om Ca+ van troponine C naar het sarcoplasmatisch reticulum te katapulteren, beide processen signaleren het begin van relaxatie. Een van de mijlpalen van de laatste jaren in het mechanisme van spiercontractie is dan ook de wetenschap dat het actieve proces van maximaal energieverbruik de relaxatie is 19,20 . Daarom kan de hartcyclus nog steeds worden verdeeld in systole en diastole zoals traditioneel sinds Harvey wordt gedaan, maar met de wetenschap dat het eerste deel van de diastole actief is, d.w.z. de isovolumetrische relaxatiefase en de snelle ventriculaire vullings- of zuigingsfase.
Bij het zuigmechanisme zijn vele krachten betrokken, waarvan sommige zeer sterk moeten zijn om het geval van Roberts et al 21 te verklaren, waarbij een schotwond in de rechterhartkamer zo’n hevige zuigkracht veroorzaakte dat het linkeratriumaanhangsel in de linkerhartkamer invagineerde. De passieve elastische terugslag van de ventriculaire helix bij het ontspannen kan van fundamenteel belang zijn, hoewel het “bloedtoevoermouw”-effect van de kransslagaders bij het vullen in de diastole en de wet van Laplace bij het verdunnen van de ventriculaire wand in de diastole en het toenemen van de straal ook een rol kunnen spelen. Onlangs heeft Torrent Guasp gesuggereerd dat het hart functioneert als een zuiger waarbij de uitwerping geschiedt door het telescoperen van de basale lus over de apexische lus (afdaling van de zuiger langs de cilinder) en de aanzuiging zou bestaan uit de verlenging-verwijding van de ventrikels door het rechttrekken van het septum dat de basale lus sleept (opstijging van de zuiger) 2 .
Maar dit gebeurt in de linker ventrikel. Waarom is er geen isovolumetrische relaxatie in de rechter ventrikel en dus geen zuiging? Het is duidelijk dat als er contractie is in de rechter hartkamer, er ontspanning moet zijn en als er ejectie is, er vulling moet zijn. Een mogelijkheid zou kunnen zijn dat de driehoekige rechterventrikel volume-druk curven twee verschillende en gelijktijdige gebeurtenissen registreren: contractie van het infundibulum en vulling van het veel grotere instroomgedeelte. Er zijn ten minste twee documenten die dit standpunt ondersteunen. March et al 21 suggereerden dat de rechterkamer uit twee onafhankelijke kamers bestaat en Pouleur et al 22 bevestigden dat de uitdrijving van de rechterkamer doorgaat wanneer de vrije wand ontspant, hetgeen Shaver et al vanuit auscultatie-oogpunt reeds hadden bepleit en als “wurging” hadden aangeduid om de vertraging van de pulmonale component van de tweede toon te verklaren.
De andere mogelijkheid zou zijn dat de lage impedantie van de pulmonale boom geen zuigfase vereist en de rechter harthelft vult door eenvoudige atrioventriculaire drukval. Voorstander van deze eenvoudiger zienswijze zou zijn dat als er geen isovolumetrische relaxatiefase is, er ook geen isovolumetrische contractiefase nodig is, die ook afwezig is in de volume-drukkrommen van Grignola et al. en in plaats daarvan optreedt bij pulmonale hypertensie, waar de volume-drukkrommen gaan lijken op die aan de linkerkant.
Is het de hoge nabelasting die een fase van isovolumetrische contractie en isovolumetrische ontspanning vereist om zuiging te veroorzaken?
Hoe dan ook, de verschillen tussen de driehoekige vorm van de rechterventrikel volume-druk curven en de kwadratische linkerventrikel volume-druk curven zijn een fascinerend en objectief feit, waarvan de hemodynamische interpretatie nadere beschouwing en studie behoeft.